백내장은 황반변성, 녹내장 등과 더불어 시력저하를 일으키는 안과 질환 중 하나로, 여전히 전 세계적으로 주요한 실명의 원인으로 알려져 있다[1]. 수술 도구나 기법의 발달로 백내장으로 인해 실명이 되는 환자는 감소하고 있으나, 인구의 고령화가 진행됨으로써 그에 따른 환자들의 비용 부담과 함께 사회 경제적 비용도 증가하는 것으로 보고되고 있다[2]. 백내장의 원인에는 선천적 요인 뿐만 아니라 후천적, 환경적 요인이 작용하며, 그 발생 위험인자로는 고령, 당뇨, 스테로이드, 외상, 그리고 자외선 노출 등이 알려져 있다[3]. 이 중에서 스테로이드 유발 백내장은 전신적으로나 점안 스테로이드를 투여 받은 환자에서 발생하는 백내장으로 정의되는데, 평균 수명의 연장과 더불어 자가 면역질환 등 장기적인 스테로이드 치료가 필요한 질환의 증가 등으로 인해 유병률이 점차 높아지는 백내장의 아형으로 보고되고 있다[4]. 스테로이드 유발 백내장은 임상적으로 중심성 후낭하혼탁의 형태로 나타나게 되는데, 이에 따라 조기에 시력 저하나 눈부심 등이 생기게 된다. 뿐만 아니라 수술 중에는 후낭파열이 생길 위험성이 높으며, 수술 후에는 후낭혼탁이 쉽게 발생하여 추가적인 비용을 발생시키게 된다[5-7]. 이런 임상적인 중요성에도 불구하고 스테로이드 유발 백내장에 대한 기전이나 예방에 대한 연구는 당뇨병성 백내장이나 연령관련 백내장보다 적은 것이 현실이다. 본 연구에서는 수정체 상피세포를 이용하여 스테로이드가 수정체 상피세포에 미치는 영향을 확인하고 이를 polymerase chain reaction (PCR) array 방법을 통해 유전자들의 발현 변화를 확인함으로써 스테로이드 유발 백내장의 병리 기전을 알아보고자 한다.
대상과 방법
세포 배양 및 처치
인간 수정체 상피(Human lens epithelial B3, HLE-B3) 세포는 American Type Culture Collection (ATCC, Rockville, MD, USA)에서 구입하였다. HLE-B3 세포를 10% fetal bovine serum (FBS) 및 1% penicillin/streptomycin (Gibco BRL, Grand Island, NY, USA)을 첨가한 Dulbecco's modified Eagle's (DMEM) 배지에서 배양하였다. 세포를 가습된 CO2 배양기에서 37°C로 유지시켰다. 대조군 세포는 덱사메타손(Sigma-Aldrich, St. Louis, MO, USA)을 처리하지 않았으며 처치군 세포는 3군(0.01, 0.1, 1 mg/mL)으로 나누어 24, 48, 72시간 동안 덱사메타손을 처리하였다.
세포 생존 분석
HLE-B3 세포를 well당 5×104 세포의 밀도로 24-well 배양 플레이트에 접종하고 24시간 동안 부착시켰다. 다음날, 세포를 24, 48, 72시간 동안 다양한 농도의 덱사메타손(0, 0.01, 0.1 및 1 mg/mL)로 처리하였다. Cell Counting Kit-8 (CCK-8, Dojindo Laboratories, Kumamoto, Japan)을 사용하여 세포 성장을 평가하였다. 10 μL의 CCK-8 용액을 각 well에 첨가하고 95% 공기 및 5% CO2의 가습 환경에서 37°C에서 1시간 동안 배양했다. 황색 formazan 염색량을 ELISA 플레이트 판독기(Molecular Devices, Sunnyvale, CA, USA)를 사용하여 450 nm에서 흡광도를 측정하였다.
세포 이동성 분석
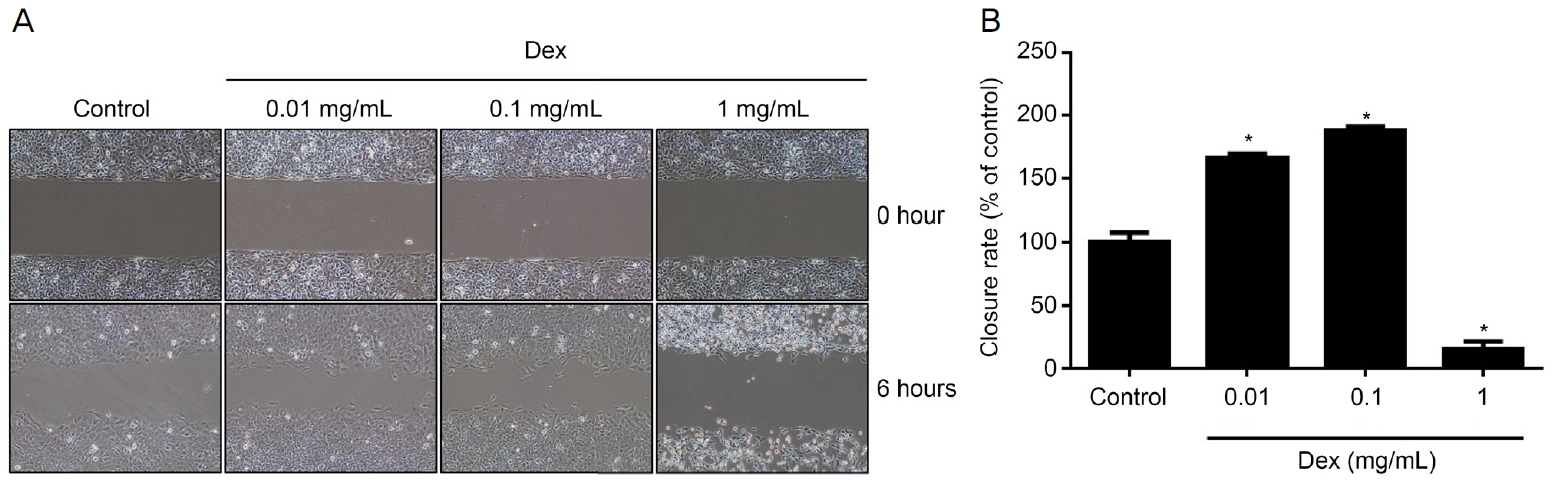
세포의 운동성은 wound healing assay chambers (ibidi GmbH, Munich, Germany)를 이용하여 측정하였다. FBS를 함유하는 DMEM에 현탁한 후, 4×104개의 HLE-B3 세포를 각각의 well에 접종하고 24시간동안 배양 플레이트에 부착시켰다. 다음 날, wound assay chambers 제거하고 현미경 이미지를 얻었다. 그 후, 세포를 덱사메타손으로 6시간 동안 처리하고 위상차현미경을 사용하여 이미지화하였다. 이미지 분석은 Nikon NIS Elements (Nikon, Tokyo, Japan) 소프트웨어를 사용하였다.
Western blot assay
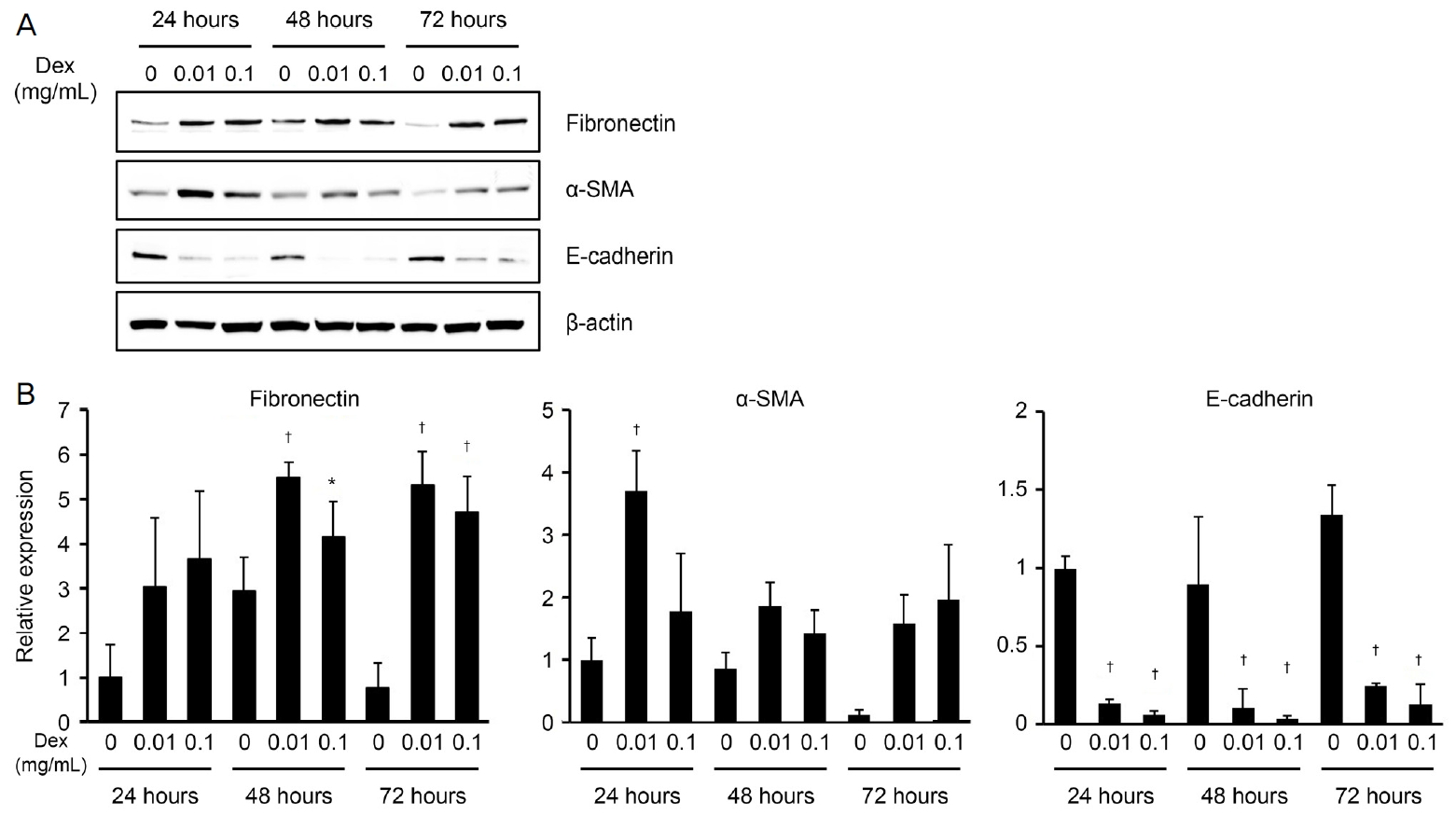
세포 추출물은 protease inhibitor와 phosphatase inhibitor cocktail (Thermo-Fisher Scientific, Waltham, MA, USA)가 첨가된 radioimmunoprecipitation assay (RIPA) lysis 완충액(Santa Cruz Biotechnology, Santa Cruz, CA, USA)를 사용하였다. 세포를 2분 동안 초음파 처리하고 4°C에서 10분 동안 12,000 g에서 원심 분리하여 불용성 세포 잔해를 제거하였다. 용해된 세포의 단백질 농도는 bicinchoninic acid (BCA) 단백질 분석 키트(Pierce, Rockford, IL, USA)로 측정하였다. 총 50 μg의 단백질을 10% sodium dodecyl sulfate polyacrylamide 겔 전기 영동으로 분리하고 nitrocellulose membrane (Millipore, Bedford, MA, USA)으로 옮겼다. 각각의 blot에 fibronectin, α-smooth muscle actin (α-SMA), E-cadherin (Santa Cruz Biotechnology) 및 β-actin (Sigma-Aldrich)의 일차 항체를 배양하였다. 이후 이차 항체로 horseradish peroxidase-conjugated anti-rabbit immunoglobulin G (IgG) 혹은 anti-mouse IgG (Cell Signaling Technology, Inc., Beverly, MA, USA)를 배양하였다. 각각의 신호는 Supersignal Chemiluminescent Substrate (Pierce)를 사용하여 시각화하였다. 이미지는 ChemiDoc Touch Imaging System (Bio-Rad, Hercules, CA, USA)으로 획득하였고 농도 측정은 Image J (NIH, Bethesda, MD, USA)를 이용하였다.
Real time (RT)-PCR array 분석
덱사메타손이 처리된 세포에서 84개의 인간 성장 인자 관련 유전자의 발현을 384개의 well (Qiagen, Valencia, CA, USA)에서 정량적 RT-PCR array를 사용하여 동시에 분석하였다(Table 1). 덱사메타손 0.1 mg/mL를 세포에 처치한 후 24, 48, 72시간 배양 후 RNeasy kit (Qiagen)를 사용하여 HLE-B3 세포에서 전체 RNA를 분리하였다. 추출된 RNA에 오염된 Genomic DNA는 RNase-Free DNase set (Qiagen)을 이용하여 제거하였다. 총 RNA (5 μg)는 RT2 First Strand Kit (Qiagen)을 사용하여 10 μL의 반응 부피의 cDNA로 변환하였다. 변환된 cDNA를 RT2 SYBR Green Mastermix (Qiagen) 및 증류수로 4,200 μL의 부피로 희석시켰다. 그 후, 10 μL의 cDNA를 인간 성장 인자 RT2 Profiler PCR Array (Qiagen)의 각 primer 세트와 제조사의 프로토콜에 따라 결합시켰다. 증폭 반응은 ViiA7 Real-Time PCR System (Applied Biosystems, Foster City, CA, USA)에서 자동화된 기준점 및 역치 주기 검출을 사용하였다. 성장 인자 관련 유전자의 상대적 발현은 4개의 내장된 housekeeping 유전자에 대해 정성화하였다. 각 그룹은 웹 기반 소프트웨어인 RT2 Profiler PCR Array Data Analysis (SABiosciences Corp., Frederick, MD, USA)를 이용하여 비교하였으며, 이 소프트웨어에서 TargetScan 알고리즘과 miRBase의 miRNome의 검색을 통해 miRNA 조절 유전자 결과를 얻었다.
결 과
HLE-B3 세포 생존에 덱사메타손의 영향
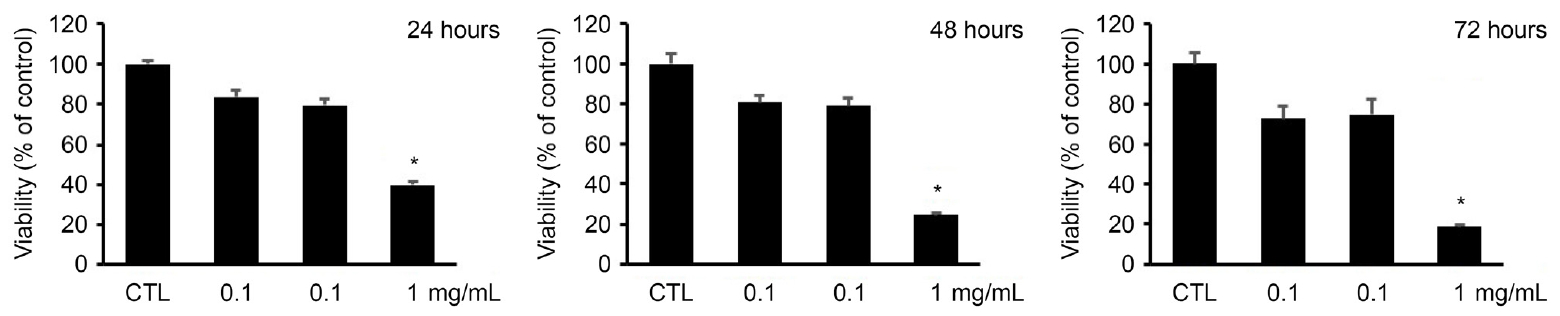
각각 다른 농도의 덱사메타손(0.01, 0.1, 1 mg/mL)에 24, 48, 72시간 노출시킨 후 대조군과 비교하였을 때 0.01, 0.1 mg/mL의 농도에서는 세포 생존율이 감소 경향을 보였으나 이는 통계적인 의미가 없었으며, 1 mg/mL의 덱사메타손에 노출 시에는 24, 48, 그리고 72시간 모두에서 세포 생존이 유의하게 감소함을 보였다(Fig. 1).
HLE-B3 세포 창상 회복에 덱사메타손의 영향
덱사메타손을 처리하지 않은 대조군과 비교하였을 때, 0.01, 0.1 mg/mL 농도의 덱사메타손에 6시간 노출 시 농도에 따른 세포 이동의 증가를 확인할 수 있었지만, 1 mg/mL 덱사메타손에 노출한 경우 대조군에 비해 세포의 이동의 감소가 의미 있게 나타났다(Fig. 2).
HLE-B3 세포의 상피-중간엽 이행에 덱사메타손의 영향
0.01과 0.1 mg/mL 농도의 덱사메타손에 24, 48, 72시간 노출시킨 후 epithelial-mesenchymal transition (EMT)에 관여하는 표지자들의 발현 변화를 확인해 본 결과, fibronectin은 덱사메타손 노출 후 48, 72시간의 모든 농도에서 대조군에 비해 증가하는 것을 확인하였으며, α-SMA는 대조군에 비해 전반적으로 증가하는 경향을 보였으나, 통계적으로는 0.01 mg/mL의 농도로 24시간 처리한 군에서만 통계학적으로 의미 있는 증가를 보였다. 반면에 상피세포의 표지자인 E-cadherin의 mRNA의 발현은 모든 노출 시간과 농도에서 대조군에 비해 의미 있게 감소하는 소견을 관찰할 수 있었다(Fig. 3).
PCR array 결과
PCR array에서 chondroitin sulfate proteoglycan (CSPG) 5가 덱사메타손 처치 후 24, 48, 72시간 모두에서 약 2배 (1.9593; 2.3975; 2.0076)로 발현 증가하였으며 24시간 노출 시 fibroblast growth factor (FGF) 11, C-fos induced growth factor (FIGF), glucose-6-phosphate isomerase (GPI), interleukin (IL) 12B, inhibin, alpha (INHA), nodal homolog (NODAL), solute carrier (SLCO1A)2와 48시간 노출 시 FGF19, INHA, NODAL, neurturin (NRTN), 72시간 노출 시 bone morphogenetic protein (BMP) 6, FGF9이 의미 있게 증가하였다(Table 2). 반면에 brain-derived neurotrophic factor (BDNF), BMP5, dickkopf homolog (DKK) 1, FGF1, FGF2, FGF5, glial cell derived neurotrophic factor (GDNF), heparin-binding EGF-like growth factor (HBEGF), IL11, inhibin, beta A (INHBA), leukemia inhibitory factor (LIF), nerve growth factor (NGF), neuregulin (NRG) 1, pleiotrophin (PTN)은 모든 노출 시간에서 발현 감소를 보였으며, IL1B는 24, 72시간 덱사메타손 노출 시 감소하고 vascular endothelial growth factor C (VEGFC)는 48시간 덱사메타손에 노출 시 감소하는 소견을 보였다(Table 3). RT2 Profiler PCR Array Data Analysis의 TargetScan 알고리즘과 miRBase의 miRNome의 검색을 통해 후보자 miRNA를 검색하여 표와 같은 결과를 얻었다(Table 4).
고 찰
본 연구에서 수정체 상피세포에서 스테로이드에 노출 시 EMT에 미치는 영향과 이를 PCR array를 이용하여 유전자들이 발현의 변화를 살펴보았다. 수정체 상피세포에 덱사메타손을 처리하면 농도 의존적으로 세포의 생존율이 감소하며, 세포의 이동과 상피-중간엽 이행 표지자의 발현 증가를 확인할 수 있었으며, PCR array에서 여러 성장인자들의 발현의 증가와 감소를 확인할 수 있었다.
장기간의 전신적 또는 국소 스테로이드의 사용은 백내장 발생을 증가시킨다고 알려져 있으며 특히 후낭하혼탁 백내장 발생에 연관성이 있다고 알려져 있다[4,5]. 이전의 연구에서 국소 스테로이드를 점안 후 약 15-30분 후 전방수에서 스테로이드가 측정이 되며, 0.1% mg/mL의 현탁액을 점안한 경우 91-120분에 최고 농도로 다다르며 이는 약 31 ng/mL 정도로 보고되고 점안 후 12시간까지 전방수에서 측정됨이 알려져 있다[8]. 또한 전신적 스테로이드 사용 후 안구 조직 내로 이동되는 농도에 대한 동물 연구에서 전신으로 투여되는 스테로이드 양의 약 0.5-1%의 스테로이드가 안구 내에서 발견되는 것으로 알려져 있고 장기간 사용할수록 더 높은 양이 발견되는 것으로 보고되었다[9]. 따라서, 류마티스 질환이나 흡입용 스테로이드를 사용하는 폐질환의 증가는 스테로이드 사용의 증가로 이어지고 이는 스테로이드 유발 백내장의 발생을 증가시키는 결과를 가져오게 된다[10,11]. 따라서 평균 수명 증가에 따른 연령 증가는 연령에 따른 백내장과 더불어 스테로이드 유발 백내장 발생률의 증가가 예상된다. 하지만 이러한 임상적 중요성의 증가에도 스테로이드 유발 백내장의 발생 병리기전에 대한 연구는 미흡한 상태이다.
스테로이드 유발 백내장의 발생 기전에 대해 수정체 글루티코르티코이드수용체의 역할에 대한 연구가 잘 알려져 있다[5,12-14]. 사실 후낭하혼탁 형태로 발생하는 스테로이드 유발 백내장에서 원래 수정체상피세포가 존재하지 않는 후낭으로 세포가 이동할 수 있도록 이동성이 증가하여야 하며, 이후에 이동한 세포에서 상피-중간엽 이행이 발생하여 백내장이 생긴다고 한다[5,13,15]. 본 연구에서도 수정체 상피세포에 독성을 유발하지 않는 스테로이드 농도(0.01, 0.1 mg/mL)에서 세포의 이동이 대조군에 비해 유의하게 증가됨을 확인할 수 있었다. 또한 스테로이드에 노출된 수정체 상피세포에서 EMT의 표지자인 fibronectin과 α-SMA를 유도하는 mRNA의 발현이 대조군과 비교하여 유의하게 증가하였으며, 상피세포의 표지자인 E-cadherin의 mRNA의 발현은 의미 있게 감소함을 확인함으로써 스테로이드에 노출된 수정체 상피세포의 EMT 증가를 확인할 수 있었다.
상피-중간엽 이행과 관련된 성장인자들의 역할에 대해 많은 연구들이 있다. Jobling and Augusteyn [4]은 후낭하혼탁 백내장에서 처음으로 안구내 성장인자의 역할을 보고하였으며, 이들 중 수정체 상피세포의 증식과 섬유세포로의 분화에 관련된 성장인자로 FGF2, IGF1, EGF, TGFβ, LEDGF, platelet derived growth factor (PDGF), BMP 등이 있다고 보고하고 있다[16-27]. 이번 연구에서 수정체 상피세포의 스테로이드 처리 후 PCR array를 통한 성장인자 분석 시 이미 알려진 성장인자 이외에도 BMP6, CSPG5, FGF9, FGF11, FGF19, FIGF, GPI, IL12B, INHA, NODAL, NRTN, SLCO1A2가 대조군에 비해 의미 있게 증가하는 성장인자로 나타났고, 반면에 BDNF, BMP5, DKK1, FGF1, FGF5, GDNF, HBEGF, IL11, INHBA, LIF, NGF, NRG1, PTN, VEGFC는 스테로이드를 처리한 수정체 상피세포에서 발현이 감소되는 것을 확인하였다.
수정체 상피세포에 덱사메타손 처리 후 발현이 증가하는 성장인자들 중 특이한 것은 NGF나 CSPG와 같은 신경유래 성장인자들이 증가하거나 감소하는 등의 발현 변화가 관찰된 것이다. 우선 NGF는 덱사메타손 처리 후에 수정체 상피세포에서 발현이 감소하는 것으로 결과가 나왔다. NGF는 이전 연구에서 수정체상피세포에서 자체적으로 발현이 되며, 실험적으로 그 발현이 감소하는 것이 당에 의한 백내장 발생에 관여한다는 보고가 있는데[28], 이는 이번 우리 연구와 일치하는 결과이다. 그리고 CSPG5는 발생 시기의 수정체 소포에서 발현이 되는데, 이때 장애가 발생시 수정체의 형성에 문제를 일으키는 것으로 보고되었으며, FGF2와 연관되어 작용하는 것으로 알려져 있다[29]. CSPG5는 neuroglycan C라고도 불리며 망막색소상피세포와 신경세포에서 주로 발견되며, 쥐에서 망막변성에 관여하는 인자로 보고되었다[29-32]. 하지만 CSPG5의 신호전달 체계의 확립이 되어 있지 않으며, 수정체에 대한 역할의 보고는 없다. 따라서 CSPG5의 역할 규명은 수정체 상피세포의 상피-중간엽 이행에 대한 이해와 스테로이드 유발 백내장의 발생 기전에 도움이 될 것으로 생각된다.
FGF는 수정체 상피세포의 증식과 섬유세포로의 분화를 촉진시키는 성장인자로 알려져 있으며[27], PDGF가 Phosphoinositide 3-kinase와 Mitogen-Activated Protein Kinase 신호전달 체계를 통해 FGF의 수정체 상피세포에 대한 역할을 촉진시키는 것으로 알려져 있다[33]. 본 연구에서 후보자 유전자로 선정된 FGF19은 아직까지 백내장 발생과 관련된 연구는 없으나 닭의 배아에서 눈 발생에 있어서 관여한다는 연구가 있으며[34,35], 제브라피시의 수정체의 발달에 FGF19이 관여하며[36], FGF19-FGFR4 신호전달 과정을 통해 수정체 형성을 한다는 보고가 있어[37], FGF19이 백내장 발생에도 관련이 있음을 추론해 볼 수 있겠다.
본 연구는 HLE-B3 세포를 이용한 연구로 일차세포가 아닌 세포계를 이용한 in vitro 실험이라는 제한점이 있으며, 이는 추후 in vivo 실험이나 수정체낭의 일차 배양을 통한 일차세포를 이용한 실험 등이 필요할 것으로 생각이 되며, 각각의 성장인자들의 역할과 신호전달체계에 대한 후속 연구가 필요하다. 결론적으로 수정체 상피세포에서 덱사메타손 노출은 세포 생존율 감소와 세포 이동과 관련된 인자들의 발현을 증가시키며, PCR array 방법은 스테로이드 유발 백내장에 관여하는 후보 물질을 발굴하고 이를 통해 스테로이드 유발 백내장의 억제 물질 발견 및 치료제 개발에 도움이 될 것으로 판단이 된다.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print